

- Introduction
It is a common belief among scientists that all organisms on our planet, living and extinct, are related to each other. This idea is captured by the famous image of the Tree of Life, a tree-like diagram representing the evolutionary relationships between living beings. According to this view, terrestrial life belongs to just one tree of life and at its base we should find the last universal common ancestor of all life on Earth, the so-called LUCA. The existence of a single phylogenetic tree has until recently never been really challenged.
Indeed, there are good reasons to believe that this is the case. If we look at familiar life, we can see that there are biochemical and molecular features common to all living things. For example, most of the biological molecules we know are made by covalent bonds of six elements: carbon, hydrogen, nitrogen, oxygen, phosphorus and sulfur (the famous CHNOPS elements). Other universal features are the presence of 5 nucleobases, pentose sugars and 20 amminoacids (all of them with an L-chirality) and the quasi-universality of the genetic code[1].
This is with regard to life as we know it. But this opens up a major problem, not only for biologists but also for astrobiologists, scientists who study the origin and distribution of life in the universe. The point is, how can we be sure that the biochemical and molecular characteristics we listed above are universal to the phenomenon of life throughout the universe, and not mere contingencies of how life developed on our planet? Which are the “signatures” of life that astrobiologists should look for on other planets? Should they assume that life elsewhere possesses the same characteristics as life here, and act accordingly?
This is exactly the N=1 problem. Having only one life sample (hence the N=1), how can we make plausible inferences about what life on other planets might be like if we have only one sample of life and do not know which characteristics are universal and which are contingent? To make a comparison, it would be like doing a cognitive psychology experiment on a single subject and expecting the results to be projectable to all human beings as well. Without terms of comparison, it is not easy to tell which characteristics of the study subject are common to all humans and which are idiosyncratic.
However, according to the philosopher of science Carol Cleland, it would be possible to start addressing the problem without finding a second sample of life on another planet. Cleland proposes to imagine the existence of a second, hidden biosphere, the result of a second abiogenesis (the arising of living things from non-organic matter) on Earth[2]. Cleland dubs this proposed second biosphere Shadow Biosphere[3]. Presumably, the organisms would differ from ours in some of the biochemical and/or molecular characteristics we presented earlier. In fact, nothing prohibits that life could have developed, for example, from amino acids with an L-chirality. According to Cleland, the organisms of this hidden biosphere should be microbes, otherwise presumably we would have already identified them[4].
But how could the finding of a shadow biosphere help us solve the N=1 problem? Having a second example of life available, we could begin to compare our biosphere with the shadow biosphere to see which features are (probably) contingent and which are necessary. For example, if the shadow biosphere organisms were also composed of CHNOPS atoms, we might already begin to think it more likely that this is a common feature of life in the universe. But if these organisms exploited novel metabolic pathways, then we could expand the number of biosignatures of interest to astrobiologists looking for life on other planets. And even if we found no organisms belonging to a shadow biosphere, according to Cleland this research could refine our use of tentative criteria in the search for life. It is to the discussion of these criteria that we now turn.
- A Kuhnian framework
Before analyzing further Cleland’s proposal, let’s pause for a moment to explore the context within which she operates. Cleland’s proposal fits within a Kuhnian perspective, where the focus of astrobiological scientific research must fall on finding bio-geo-chemical anomalies resisting attempts to be categorized as biological or non-biological and that might suggest the presence of life at a particular location. Cleland starts by analyzing some cases in which adherence to overly geocentric criteria of what life is has hindered further research. In particular, she presents the case study of NASA’s Viking missions to Mars[5]. One of the three experiments (the “Labeled Release” (LR) experiment), performed by the landers in 1976 sparked a lot of controversies. In short, a sample of Martian soil was inoculated with a nutrient solution, radioactively labeled with 14C, an isotope metabolizable by terrestrial bacteria (the experiment was therefore designed using life on earth as a model). Unfortunately, the results of the experiment were ambiguous, as at the same time the soil sample behaved both in ways compatible with the presence of terrestrial-like organisms and in manners that were difficult to explain, even taking into account abiotic phenomena. The PI’s who designed the experiment claimed to have found life because the recorded activities were compatible with those of terrestrial microbes, while the official NASA position is that they did not find life, since the metabolizing activity registered was in some respects completely different from terrestrial life. Both sides of the controversy failed to recognize that the results of the experiment were truly ambiguous and needed further research.
To avoid impasses like that of the Viking mission, she recommends using tentative criteria instead of defining criteria to search for extraterrestrial life, that is, criteria open to review on the basis of empirical or theoretical advances. According to Cleland, this is a strategy that would allow us to flexibly continue astrobiological research even in a scenario like ours where there is no accepted definition of life. The idea is that this is also the most promising strategy for searching for life forms belonging to the Shadow Biosphere.
- Criticism of the Shadow Biosphere
There has been a lot of criticism of the possibility of a shadow biosphere. As might be expected, critics question why, if a shadow biosphere really exists, it has not yet been identified. In the words of Charles S. Cockell, one of the prominent astrobiologists: «I think it is very unlikely that after 300 years of microbiology we would not have detected such organisms despite the fact that they are supposed to have a different biochemistry from the kind we know today»[6].
Cleland can easily answer criticisms like the one raised by Cockell. Indeed, our microscopy, cultivation and metagenomic methods, usually used to detect and study microscopic organisms, are either too fine-graned or coarse-grained to identify shadow organisms[7].
In our opinion, it is possible to advance another response to this criticism. It should be noted that we still know only a tiny fraction of the microscopic life on Earth, even taking into consideration only our biosphere[8]. An example, very recent and remarkable, is the discovery of a new supergroup (a taxonomic category of high-level recently introduced, above kingdoms) of predatory microbial eukaryotes, called provora[9]. Moreover, in 2010 a NASA-funded astrobiology research group announced an even more shocking discovery: bacteria capable of replacing phosphorus with arsenic in their DNA and proteins[10]. Understandably, this announcement sparked much controversy. Ultimately, it seems that these bacteria do not replace phosphorus with arsenic, but simply tolerate environments very rich in arsenic[11].
Clearly, discoveries such as these expand our knowledge of life, but within the current sample. That is, although they are organisms that have novel characteristics or lead us to revise our taxonomic classifications, they still share the same common ancestor of all other organisms in our biosphere, not allowing us to overcome the N=1 problem we talked about before. If nothing else, they underline how little we know about the microorganisms that inhabit our planet.
A more serious problem for the shadow biosphere is lurking in Cleland’s response to another criticism. Cleland refers to our microbial life to infer what the likely characteristics of shadow biosphere life might be. If it is quite natural to think that these organisms are microbial (otherwise we would have almost certainly noticed them), the same cannot be said for more precise characteristics that Cleland attributes to it. For example, one of the most frequent objections to the possibility that a shadow biosphere exists at present is that organisms belonging to our biosphere would have eradicated any other form of life in a fierce Darwinian competition for environmental resources. Cleland responds to this criticism by stating that in the microbial world collaboration, rather than competition, is the rule, even between organisms of different domains such as bacteria and archaea, so we should expect that the same should also happen between different biosphere organisms. This answer is problematic though, because it assumes the shadow biosphere microbes look like “ours” and are equally collaborative. But it is difficult to say whether being collaborative is a necessary trait of all microbes or contingent, and we are therefore in this ironic situation where we cannot deduce whether or not this characteristic is present in the microbes of the shadow biosphere as long as we are limited to just one example of life. The N=1 problem strikes back. The same goes for many other features.
Furthermore, Cleland does not consider a number of conceptual distinctions and issues related to its definition of the shadow biosphere. She simply speaks of a second abiogenesis, presumably occurring at the same time as the first, whose organisms would still be present today. But, as Davies and Lineweaver[12] point out, numerous alternative scenarios are possible. For example, a possibility is that life on planet Earth had only one origin, but then it was so differentiated, from the biochemical and molecular point of view, as to seem to belong to two distinct abiogenesis. Being faced with organisms radically different from the molecular and biochemical point of view, how would it be possible to understand if they are two different abiogenesis or a single abiogenesis (a single tree of life therefore) with two branches at the base that diverge in a radical way? Perhaps two different genetic codes could evolve from a simpler code, but what about, for example, organisms with opposing chirality evolved from an achiral ancestor? Unfortunately, having no idea of what characteristics of life are necessary and what contingencies it is difficult to go beyond mere speculation.
- The history of our astronomical context
As seen above, the issue with the shadow biosphere hypothesis concerns the lack of evidence about biochemically different life forms. Consequently, it is fair to ask the question: if the shadow biosphere existed or still exists, why have we not yet found such life forms? This question is certainly problematic, although the N=1 problem does not allow for the claim that the peculiar characteristics of terrestrial life are universal. However, considering the history of the Earth and our astronomical context, some hypotheses can be pointed out.
Because of the N=1 problem, the habitability of a planet is established in relation to the degree of similarity to Earth, a rocky planet in the Solar System that can therefore be considered as an astronomical context suitable for life. However, the past of our planetary system shows epochs with very different scenarios. Long ago our planet was not suitable for life at all. The history starts 4.56 Ga ago, with the beginning of a process that can be described through the solar nebula hypothesis, attributed to Emanuel Swedenborg in 1734[13] and later revised independently by Kant and Laplace[14]. This model was revised in 1969, when Russian astronomer Viktor Sergeevič Safronov advanced the planetesimal hypothesis for the formation of planets[15].
According to this theory, planets are a byproduct of the phenomenon of star birth, the result beginning with the gravitational collapse of a molecular cloud. A much more chaotic environment than what we observe today. The law of conservation of angular momentum causes matter to orbit in the same direction as the star, forming the so-called protoplanetary disk. Subsequently, the solar wind of the protostar pushes light elements (such as hydrogen) outward and chemically differentiates the inner planetary system from the outer one. According to this hypothesis, protoplanets form in about 5 million years through electrostatic and gravitational interactions, transforming dust and gas into planetesimals and, at the end, planets[16].
This process leads to the formation of gas giants like Jupiter or icy giants like Neptune beyond the “snow line”, while rocky planets like Earth, Venus and Mars formed in the inner Solar System. Before reaching its current configuration, the proto-Earth underwent numerous impacts including the one with Theia, a Mars-sized object[17]. According to this model, the collision caused the formation of the Moon, creating the Earth-Moon system we can observe today. During this phase, our planet was still far from being a hospitable place for life. However, the rate of impacts continued to decline and, according to some evidence, 4.2 Ga ago our planet was characterized by the presence of water in the liquid state and thus oceans. Although there is no irrefutable evidence for the presence of life during this epoch, some studies reveal that life may have colonized some ecological niches at least 3.95 Ga ago[18]. Thus, the second part of the Hadean may have been an ideal window for abiogenesis and the formation of early terrestrial life forms as well as the shadow biosphere.
Then, the quiet period after the storm of meteors and collisions with other planetesimals ended between 4.1 and 3.9 Ga ago. Indeed, during this timeframe the rate of impacts with asteroids, comets and other celestial bodies from the outer part of the Solar System increased. This phenomenon is known as the Late Heavy Bombardment Event and was caused by the gravitational interaction between Jupiter and Saturn[19]. This phenomenon pushed several celestial bodies toward the inner Solar System, involving all the inner planets and their satellites. If during that epoch the Earth harboured both familiar life and the shadow biosphere, the Late Heavy Bombardment Event profoundly changed the fate of the first living things on Earth, exerting a very strong selective pressure, a great filter for familiar life and, maybe, for the shadow biosphere.
As can be seen, the apparent harmony and stability of the Solar System today is the result of a chaotic and destructive past in which even a planet as suitable for life as Earth underwent numerous environmental and geological changes, which can be considered as catastrophic events. Thus, one of the possible hypotheses regarding the shadow biosphere could be: What if one of these drastic changes was the cause of the absence of the shadow biosphere?
- The Impact-Bottleneck hypothesis
The Late Heavy Bombardment Event may have been one of the first catastrophic events to have strongly affected the evolution of familiar life. Indeed, the increase of the impact rate caused major environmental changes as the increase in the planet’s temperature and volcanic activity. During this period, many ecological niches were probably destroyed, and life faced the difficult challenge of surviving the new planetary conditions. According to the Impact-Bottleneck hypothesis only life forms capable of surviving high temperatures (at least, extremotolerant) were able to adapt to these conditions[20]. In this view, the shadow biosphere would have failed in this endeavour, either going extinct or surviving in ecological niches that we cannot understand because of the N=1 problem. One possible evidence that can confirm this scenario concerns the dating of LUCA by the molecular clock. Indeed, estimates of the life span of the common ancestor of familiar life coincide with the Late Heavy Bombardment Event period (~ 3.9 Ga ago)[21]. Thus, LUCA may have been the only one among multiple possible phylogenetic tree roots that was able to survive. Alternatively, following Davies’ perspective, LUCA was one of different bacterial communities that survived the new planetary conditions[22]. However, bottleneck resistance does not imply that LUCA had to be an extremophile capable of surviving at high temperatures (thermophilic or hyperthermophilic), although hot birth models of life suggest that LUCA may have developed these adaptations. It may have been extremotolerant life forms able to survive and reproduce, generating the living things with the right adaptations to proliferate. Thus, if this hypothesis is a plausible scenario, it is necessary for older life forms in our phylogenetic tree to have developed these adaptations. According to current data, the life forms closest to the root of the tree of life (and to LUCA) are classifiable as thermophiles or hyperthermophiles, a factor that can corroborate this hypothesis, although not conclusively.
The fact that LUCA did not necessarily have to be a thermophilic or hyperthermophilic organism could be supported by an additional factor related to the Late Heavy Bombardment Event. Earlier, we emphasized the catastrophic aspect of this period, however, there is an additional factor to consider. Indeed, during a 4.5 Ga long history, the Earth underwent numerous impacts, and the most recent ones have created new environments that are potentially suitable for familiar life. Impact craters are exposed to perturbations and rain and, by collecting rainwater, they create ideal environments for life[23]. In addition, rocks from the outer Solar System possess a chemical composition that includes light elements such as hydrogen and, by reaching Earth, may have caused a local chemical enrichment. Another very important aspect concerns the changes that rocks undergo when they pass through Earth’s atmosphere. Indeed, this is a common phenomenon that can be understood by observing the shooting stars. When an asteroid passes through the atmosphere, it begins to burn, becoming visible to the naked eye. The asteroids involved in the Late Heavy Bombardment Event also underwent a similar process, which involves a potentially important change for life forms. In fact, these rocks become porous, and the pores may be an ideal environment for familiar life forms. In fact, many meteorites with these characteristics have undergone colonization by microbes, which are then able to proliferate in these niches. So, the Late Heavy Bombardment Event may have taken on the role of a large filter for its destructive potential but, at the same time, provided a series of ecological niches for LUCA.
Thus, in the Impact-Bottleneck hypothesis scenario, the shadow biosphere might have undergone the following endings:
- Total annihilation of the shadow biosphere.
- A mass extinction that greatly reduced the chances of survival of the shadow biosphere, causing it to survive in ecological niches that we cannot hypothesize.
Of course, by itself the Impact-Bottleneck hypothesis is not designed to justify the absence of the shadow biosphere. However, the scenario told from the aftermath of the Late Heavy Bombardment Event teaches how much this contingent event affected the future of life on Earth. Even if the shadow biosphere never existed, the selective pressure exerted during this period was one of the first challenges that family life faced. A contingency that could perhaps be a peculiarity of Earth’s history and thus of human history as well.
- Strange world in our astronomical context
The Impact-Bottleneck hypothesis can be used to justify the absence of the shadow biosphere, although the argument concerns the evolution of familiar life and remains valid even if its peculiar characteristics were to be universal. Only possible future research could provide new data and, maybe, proof of the existence of the shadow biosphere on Earth. However, our planet is not the only target for the search for biochemically diverse life forms. Indeed, there are several celestial bodies in the Solar System that may have hosted, or still host, extraterrestrial life forms. The first candidate is certainly Mars, which, until about 3.5 Gy ago, possessed oceans and a potentially habitable environment. However, the red planet appears today as a barren world. Perhaps one day we would find evidence of Martian life, if it ever existed, although Mars is not the only target for the search for extraterrestrial life in our planetary system.
Indeed, other celestial bodies could have environments hypothetically suitable for familiar life or, alternatively, for biochemically different living things. Unlike Mars, these celestial bodies are not planets but satellites. They are Europa (one of Jupiter’s satellites) Enceladus and Titan (two of Saturn’s satellites), worlds very different from our planet that orbit beyond the ice line, and thus far from the habitable zone (also called the Goldilock zone), studied through space probes that can collect data and material to understand their chemical composition. These worlds may be capable of harbouring life today, a hypothesis that rests on the assumption that at least microbial life is not unlikely.
Europa is smaller than our Moon and is tidally locked to Jupiter. This satellite shows a surface composed of ice enclosing an ocean, while the interior of the planet is composed of silicates and probably a metallic core. Being outside the Goldilock zone of the Solar System, Europa is not in a zone suitable for life as we know it. However, the surface ice layer, about 30 km long, protects the planet’s interior from UV radiation and makes the environment potentially suitable for life. According to current data, Europa’s ocean could be 100 km deep, with twice the volume of oceans on Earth. In addition, this satellite’s ocean could contain o or other kinds of salts, compounds that can influence water activity. One of the most widely accepted hypotheses for life on Europa involves the presence of methanogenic life forms, although the most optimistic estimates in this regard estimate much less microbial activity than in Earth’s oceans, that is, on the order of of steady-state biomass[24]. In a sense, this world may be suitable for life as we know it rather than the shadow biosphere[25].
Enceladus also shows an icy surface enclosing a salty ocean that is like environments on Earth characterized by high microbial activity, soda lakes. The Cassini probe’s analysis of the satellite’s chemical composition is done by collecting material from plumes, geyser-like features that emit some of the oceans’ contents outward. This study advances the conclusion that Enceladus contains all the CHNOPS elements. Again, the environment may be suitable for life as we know it, although the possibility that biochemically different life forms may have developed cannot be ruled out[26].
In contrast, Titan is not an icy moon, and its chemical composition is different. Thus, it could be an ideal target to test the hypothesis that biochemically different life forms exist. Titan is Saturn’s largest satellite and the only moon in the Solar System with a substantial atmosphere and surface pressure greater than that on Earth. The atmosphere is composed of 98.4 percent nitrogen, 1.4 percent methane and 0.2 percent hydrogen. The chemistry of Titan’s atmosphere exhibits the photolysis of methane (), which is converted to hydrocarbons by UV radiation, causing rain of ethane and other complex hydrocarbons and eventually forming lakes of methane and ethane. As can be seen, it can be argued that this world is not at all suitable for familiar life. However, it may be an interesting target for understanding whether life forms with different metabolic pathways can proliferate in these environments. Eventually, the discovery of such life forms would demonstrate that the characteristics of life as we know it are not universal, opening the possibility that Earth may have hosted what we now hypothetically call a shadow biosphere. Indeed, on Titan there is no water in the liquid state (at least not on the surface) but lakes of methane and ethane that assume the role of solvent. Some speculation suggests that hypothetical life forms could use alternative metabolic pathways by coupling compounds like ethylene to hydrogen. In this way they would be able to derive the energy they need to survive[27].
These strange worlds, more or less different from our planet, could harbour life and thus be a test case for the hypothesis of the universality of the characteristics of familiar life, that is related to the concept of shadow biosphere. In any case, should we be able to obtain evidence of biochemically different life forms, will we be able to recognize them?
- Conclusions
The last question represents one of the critical issues of Astrobiology and it is not exclusively concerned with possible biochemically different life forms. Indeed, in Life on Enceladus? It depends on its origins, L.M. Barge and L.E. Rodriguez showed through simulations that hydrothermal activity in the satellite’s ocean could support the presence of methanogenic life forms. However, simulations are compatible with different chemical processes, which can assume both a biogenic and abiotic nature. Assuming a high probability of abiogenesis on Enceladus, in a Bayesian framework it is possible to state that the process involving life forms is more probable[28]. However, this result is based on the implicit assumption that abiogenesis is probable. Thus, this work shows a very common difficulty, which also includes the precise dating of the presence of the earliest life forms on Earth through the study of microfossils: there are situations where the evidence is compatible with both biological and abiotic processes. This fact introduces a very important epistemological problem for the search for extraterrestrial life, which is therefore exposed to a false positive rate difficult to quantify. So, the N=1 problem is not the only source of uncertainty. However, the data collected through probes regarding the satellites described above open interesting windows that in the future could also provide a definitive answer to the question about the universality of the model of life we know.
Reasoning about the shadow biosphere hypothesis, in relation to the N=1 problem, led us toward different scenarios that sometimes amplified the issue, as you can see in the picture below.
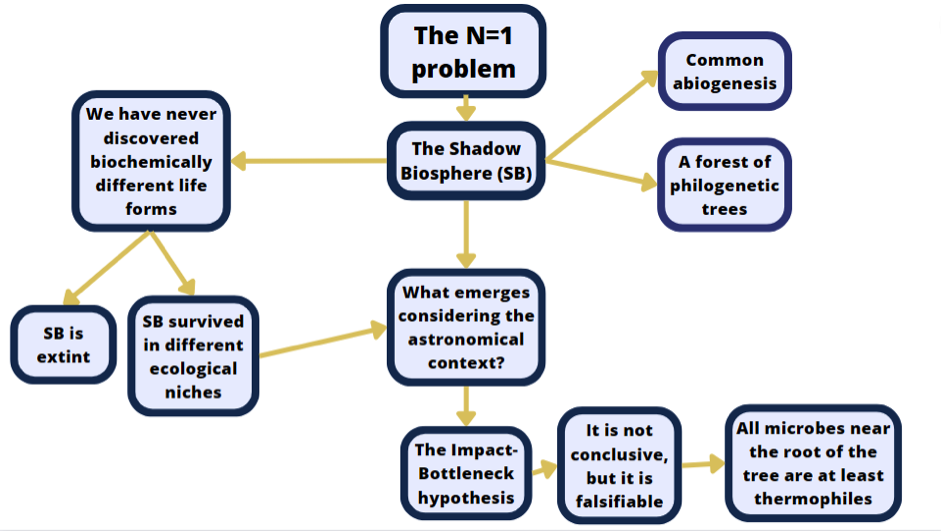
However, by referring to the history of our planet, we advance hypotheses that justify the absence of the shadow biosphere on Earth. In this sense, the Impact-Bottleneck hypothesis could be a valid justification. This attempt stems from the assumption that we want to analyze the shadow biosphere hypothesis by including it in a context that encompasses events peculiar to our astronomical environment, and thus to Earth. Finally, by focusing on the icy moons and Titan, we have shown possible targets that may allow, in the future, to test the possibility that the peculiar features of familiar life are universal. Therefore, research over the next few years on these environments will offer crucial information on whether the model of life we know is universal, thus increasing the validity of this assumption for the search for extraterrestrial life forms beyond the Solar System.
[1] C. Cockell, Astrobiology. Understand life in the Universe, Wiley Blackwell, Oxford 2020.
[2] C.E. Cleland, Epistemological issues in the study of microbial life: alternative terran biospheres?, in «Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences», 38, 4, 2007, pp. 847-861; Id., The Quest for a Universal Theory of Life: Searching for Life as we don't know it (Vol. 11), Cambridge University Press, Cambridge 2019; Id., S.D. Copley, The possibility of alternative microbial life on Earth, in «International Journal of Astrobiology», 4, 3-4, 2005, pp. 165-173; P.C.W. Davies, S.A. Benner, C.E. Cleland, C.H. Lineweaver, C.P. McKay, F. Wolfe-Simon, Signatures of a shadow biosphere, in «Astrobiology», 9, 2, 2009, pp. 241-249.
[3] Lowercase from now on.
[4] A similar perspective can also be found in E.O. Wilson, The Meaning of Human Existence, W.W. Norton & Company, New York 2014.
[5] A.C. Schuerger, B.C. Clark, Viking biology experiments: lessons learned and the role of ecology in future Mars life-detection experiments, in «Space Sci. Rev.», 135, 2008, pp. 233–243.
[6] Cited in R. McKie, ‘Shadow biosphere’ theory gaining scientific support, in «The Observer», 2013.
[7] C.E. Cleland, The Quest for a Universal Theory of Life: Searching for Life as we don't know it (Vol. 11), cit., pp. 206-211.
[8] As Cleland rightly emphasizes, it seems that just 1% of all the species of bacteria and archaea can be cultivated in the laboratory; N.R. Pace, A molecular view of microbial diversity and the biosphere, in «Science», 276, 1997, pp. 734–740.
[9] G. Lax, Y. Eglit, L. Eme et al, Hemimastigophora is a novel supra-kingdom-level lineage of eukaryotes, in «Nature», 564, 2018, pp. 410–414.
[10] D. Brown, C. Weselby, NASA-funded research discovers life built with toxic chemical, 2 December 2010 http://www.nasa.gov/topics/universe/features/astrobiology_toxic_chemical.html . Accessed November 2023.
[11] E.C. Parke, What could arsenic bacteria teach us about life?, in «Biol Philos», 28, 2013, pp. 205–218, reconstruct the long diatribe well.
[12] C.E. Cleland, S.D. Copley, The possibility of alternative microbial life on Earth, in «International Journal of Astrobiology», 4, 3-4, 2005, pp. 165-173.
[13] E. Swedenborg, Opera Philosophica et Mineralia, volume I, Sumptibus, Friderici Hekelii, 1734.
[14] P.S. Laplace, Traité de mécanique céleste, Charles Crapelet, Paris 1802.
[15] V.S. Safronov, Evolution of the Protoplanetary Cloud and Formation of the Earth and the Planets, in «NASA TT F-677», vol. 677, 1972.
[16] C. Cockell, Astrobiology. Understand life in the Universe, cit.
[17] Q. Yuan, Q., M. Li, S.J. Desch et al., Moon-forming impactor as a source of Earth’s basal mantle anomalies, in «Nature» vol. 623, 2023, pp. 95–99.
[18] T. Tashiro, A. Ishida, M. Hori et al., Early trace of life from 3.95 Ga sedimentary rocks in Labrador, Canada, in «Nature», vol. 549, 2017, pp. 516–518.
[19] A. Mann, Bashing holes in the tale of Earth’s troubled youth, in «Nature», vol. 553, 2018, pp. 393-395.
[20] M. Gogarten-Boekels, E. Hilario, J.P.Gogarten, The effects of heavy meteorite bombardment on the early evolution – the emergence of the three domains of life, in «Origins of Life and Evolution of Biospheres», vol. 25, 1–3, 1995, pp. 251–264.
[21] C. Cockell, Astrobiology. Understand life in the Universe, cit.
[22] G.P. Morales, L. Delaye, Was LUCA a Hyperthermophilic Prokaryote? The Impact-Bottleneck Hypothesis Revisited, in V. Souza, A. Segura, J. Foster, (eds), Astrobiology and Cuatro Ciénegas Basin as an Analog of Early Earth, Springer, Berlin 2020.
[23] G.R. Osinski, C.S. Cockell, A. Pontefract, H.M. Sapers, The Role of Meteorite Impacts in the Origin of Life, in «Astrobiology», Sep 2020, pp. 1121-1149.
[24] C. Cockell, Astrobiology. Understand life in the Universe, cit.
[25] Ibid.
[26] Ibid.
[27] Ibid.
[28] L. M. Barge, L.E. Rodriguez, Life on Enceladus? It depends on its origins, in «Nature Astronomy», vol. 5, 2021, pp. 740–741.





