

- Introduction: science and philosophy in 19th century Britain
Isaac Newton’s universal gravitational theory and its further corroboration gave induction, the inferential process that allows the building of hypotheses on a previous number of observable cases, a special place among the methods in science. Newton advocated his employment of observation as the securest path to truth, and the following decades were instrumental in confirming the strength of Newtonian physics. This would have had a lasting influence on 19th century philosophers, notwithstanding Hume’s critique of such process of reasoning[1]. William Whewell’s consilience of inductions, and John Herschel’s search after nature’s true causes, shaped the way science was looked at. According to the former, multiple evidential strands, gathered inductively, could provide strong support for a theory; Herschel strengthened this idea by assuming that scientists should admit in their theorising only those causes that could be ascertained, contributing to diminishing the influence of classical metaphysics over science[2].
In this context, physics and astronomy were the first among the sciences for their experimental character and their predictive success. Indeed, successful, and unexpected confirmation of predictions were looked at as being superior to mere explanation of known evidence, a factor that caused Darwin’s work to almost go unnoticed by major philosophers of the time. Another factor influencing the theory’s reception was the essentialist and typological outlook of biological organisms, as well as the teleology believed to be intrinsic in the natural world. Johann Wolfgang Goethe (1749-1832), Etienne Geoffroy Saint-Hilaire (1772-1844) and Richard Owen (1804-1892) understood organisms and their biological structure as many variations on the same platonic theme (an ideal archetype) created by God, who had made nature harmonious and purposeful[3]. By eliminating teleological explanations from our understanding of the biological world, Darwin understood that evolutionary biology could over time become a science and, I argue, the following theses were central to the theory’s establishment as a respectable area of enquiry.
- Strengthening the theory: the employment of a diversified methodology
Throughout his work, Darwin lays the foundation of modern biology emancipating its methodologies from those of physicists. Notwithstanding the scarce reception that Darwin’s work had among his contemporaries, his methods have proven robust. Darwin, besides performing a considerable number of experiments to support his theory, relied on inferential techniques that did not conform to the experimental methods adopted by sciences such as chemistry and physics. Those methods, nonetheless, allowed hypothesis building through the gathering of scattered and diverse traces through the fossil and geological record: these investigations are characteristic of the historical sciences such as geology, palaeontology, archaeology, and astrophysics.
Carol Cleland has vindicated the work of historical scientists by arguing that reconstructing past events, explaining rather than (just) predicting, is a successful enterprise[4]. By discussing the methodological flaws of purely inductive and falsificationist strategies, which deliver a simplified picture of how science works, Cleland has pointed to the study of «smoking guns» (evidential traces pointing to the same direction) as being an essential aspect of the historical sciences. Events such as geological upheavals and extinctions leave behind a high number of traces, a low number of which is sufficient to infer what happened. For instance, volcanic eruptions leave behind a significant amount of solidified lava, carbonised animals and plants, chemical elements in the ground, and so on, although scientists need only a small sample of the total evidence produced to ascertain the occurrence of an eruption.
Alan Currie has a different, albeit complementary, perspective on the topic, arguing that we should be optimistic about the success of the historical sciences, even if the distinction between experimental and historical disciplines is nowadays more blurred[5]. Specifically, Currie, following Thomas Kuhn[6], has emphasised the contextual nature of scientific hypotheses, which are originated and developed in what he terms epistemic situations. When dealing with past traces such as extinction events, there is no reason to adopt an a priori pessimistic stance on the matter. As the author rightly emphasises, there are several reasons why our inferential reasoning about the past can be successful. Firstly, scientists do not only link past to present events, but they do also relate past events among themselves, and this allows to confer greater robustness to the hypothesis under investigation. Secondly, the relevance of some piece of evidence can change over time throughout a combination of new developments in methods, theories and empirical discoveries: this fact should make scientists adopt an optimistic perspective on scientific practice, which might at times result in theoretical stagnation. Lastly, scientists are methodological omnivores, in that they successfully co-opt methods from other sciences to apply them in a fruitful manner and this enhances the theoretical flexibility and robustness of the historical enquiries[7].
In this context, several hypotheses can be put to test at the same time, and the best explanation chosen according to the greater or lesser coherence of the hypotheses to background knowledge, the context in which the evidence has been gathered and the soundness of the methods of analysis employed. By combining laboratory experiments, evidential smoking guns and (nowadays) computer simulations, it is possible to boot-strap across different and independent strands of evidence conferring strength to a given hypothesis.
Before moving to a discussion of Darwin’s approach, it is worth mentioning a different perspective on the work of historical scientists, whose main contrast with Cleland is on the emphasis of theory-ladenness of the employment of evidential traces. Bonnin has drawn on the work of Stephen Toulmin (2003)[8] in order to reconstruct the inferential processes that led to the formulation and further elaboration of the «Archezoa hypothesis»[9].
Toulmin schemas, which employ a diagrammatic representation of inferential reasoning, are structured as follows. Some ‘brute’ facts are supported by a series of warrants, that can be further backed up or refuted as the theory develops in the light of new evidential and theoretical contributions. These schemes allow all the knowledge on a hypothesis at time t to be visualised clearly, in such a way to make easier to spot flaws in the inferential process as well as to highlight the presence of ‘bold’ claims and assess their relevance for future research.
In its initial formulation, Archezoa grouped four major phyla: Archamobae, Metamonada, Microspora and Parabasalia. As new empirical discoveries were made, the hypothesis was revised several times and from A (fig below; after Bonnin 2019) evolutionary biologists rejected the membership of the four phyla to Archezoa, radically transforming the original hypothesis[10].
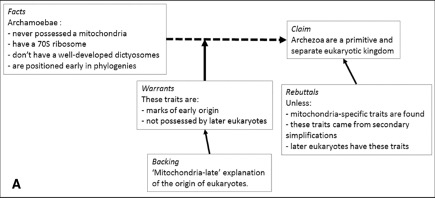
I believe that the research above has been fundamental in showing the strengths of the historical sciences which, employed along the experimental ones, can boost the epistemic status of evolutionary biology. In its rudimental form, this process was started by Darwin in The origin, by employing historical and experimental methods, and showing how the study of evolution could benefit from intertwining diverse methods. On one side, the breeding of pigeons and the analysis of the races formed[11], the study of migration patterns through the resistance of seeds in different aqueous media and the presence and absence of features allowing the transportation by clinging to animal coat[12], the co-evolution of plant-insect relationship, are clear examples of the employment of experimental techniques.
Indeed, those reported by Darwin were not simple observations. In many cases he performed experiments. He designed (a) what to investigate, (b) set up the experimental conditions, (c) with which he regularly interfered in order to monitor the consequences of such changes. Moreover (d) he reported the results of the tests periodically and made them replicable.
Concerning the means of dispersal of plant seeds, Darwin reports differential germination of some of them after being immersed in sea water for several intervals of time. In experimenting, Darwin selected the size of the seeds, registered the buoyancy of the larger seeds and, by adding these results to estimations of the speed oceanic currents, calculated for how much time and in what conditions the seeds could retain their power to germinate. To these experiments, Darwin added the observations that the seeds could be accidentally transported through drifting timbers or attached to the claws of birds retaining some dirt. He concluded that all these data put together could explain the biogeographical distribution of some plants[13].
On the other, the study of past and present disposition of the continental plaques, the analysis of the fossil sequences[14], all required the formation of hypotheses arrived at through the study of smoking guns and inferences that allowed Darwin, over many places of the work, to infer that his were better explanations than those provided by creationists, on the ground that the latter lacked logical and evidential parsimony.
For instance, from the analysis of the paleontological record he derived generalisations which he employed in the formation of his theory of organic evolution. More specifically, he claimed that (1) new species have appeared slowly; (2) species of different genera and classes underwent differential rates of change; (3) in members of the same class, given equal time, the amount of change should be the same; (4) extinct species should never reappear; (5) groups of species follow the same rules in appearance and disappearance as do single species[15].
From these instances it is clear how Darwin was able to intertwine several methods that allowed him to complement the experimental dimension of evolutionary biology, gleaned up from observations and experiments, with inferential processes characteristic of the historical sciences. In such a way he was able to explain a range of facts, unaccounted for by other methods.
- Interweaving multiple strands of evidence
Carl Hempel has argued that for a theory, the more varied the evidence is, the better it supports it[16]. In discussing Snell’s law[17], he shows that by setting different experiments in which the nature of the media or the angle of incidence are varied while the other component is kept constant, allows greater evidential support. This occurs because changing the conditions of the experiment provides a test for counterfactuals and check across how many domains the hypothesis holds. Recent work in a Bayesian perspective, has shed new light on the thesis that the more diverse the evidence, the stronger the confirmation of a theory is[18]. Although the validity of such a thesis cannot be taken to be universal, there are still cases in which it can be applied and employed successfully as a heuristic principle.
This procedure has been followed by Darwin in The origin, where he brings multiple strands of evidence to support the theory. By referring to (1) the paleontological record, Darwin infers that the diversity of existing species is due to a divergence of organic forms over time, and he predicts that the older a common ancestor is, the greater is the number of descendants it links. Darwin predicts that different species and genera will undergo different selective pressures, and that adaptations and extinctions will be varied too across different lineages. The overall picture delivered by the paleontological record supports the explicative nucleus of common descent and natural selection. Darwin, moreover, openly rejects the hypothesis of independent creations, in that it would require many non-parsimonious assumptions. Even if Darwin’s theory and creationist hypotheses are both empirically adequate[19], the theory of natural selection coupled with common descent combines a greater number of evidential strands and explained those difficulties encountered by natural theologians. For example, the imperfection of the record was explained away through taphonomic processes and cases of convergent or divergent evolution better explain common descent or adaptation to the local conditions than does the invocation of independent creations, a logically more expensive move.
It is the discussion of (2) the biogeographical record which further strengthens the support to his theory. By performing a variety of experiments on the distribution of plant species, their survival in salty water across different time spans and their greater or lesser likelihood of being transported by animals or physical agents over natural barriers, Darwin predicts and explains that:
A - Species living in different parts of the world subject to similar environmental conditions do not display similar adaptations[20].
B - Across within one determined area there will be greater affinity of the species than across different geographical spaces[21].
C - Migratory patterns explain the adaptation of foreign species, which gradually bring to the extinction of the indigenous ones[22].
Again, biogeographical patterns are better explained by Darwin’s theory than by independent creations. By interweaving the current distribution of organisms with a theory of migrations, and by comparing the biogeography of extant species to the fossil record, Darwin can show how these patterns can be explained by natural processes. Moreover, climatic changes have determined a change in the constitution of habitats and prompted organisms to migrate across vast areas. When for example, artic species vastly distributed are forced to migrate further north, they will be hindered by hotter climates to return to their original habitats and will be confined on mountainous habitats[23]. Darwin then argues that common descent and the inheritance of the modifications explain the biogeographical record.
Biogeography and palaeontology are complemented by (3) embryological studies. On Darwin’s developmental insights, some of which have now proved wrong, embryology is the key to understanding homologies and analogies across organisms. Species related by a common progenitor would show high similarity in their embryos, that tend to differentiate later according to the law of correspondence[24]; at the same time, embryonal differences do not show that some organisms are not related, because the stages might have been compressed over phylogenetic development. In his view, vestigial organs and those that contribute less to the vital functions of plants and animals are the best key to taxonomic classification, since they have been modified less by natural selection and hint at common descent[25]. In addition, Darwin rules out progress in the organisation of phylogenetic and ontogenetic development, and his studies on cirripeds show that reversals of form are possible. The study of embryological and morphological characters is pivotal in showing how the unity of type, i.e. common descent, and the action of the conditions of existence, which lead to the production of analogies, closely interact and provide a unifying perspective on the ontogenetic and phylogenetic development of individuals and groups, de facto explaining the diversity left unaccounted for by creationist theories[26].
To recapitulate, Darwin’s theory receives support by bringing multiple strands of evidence, independent of one another. Hypotheses of independent creations would require a degree of ad hocness, i.e. purposeful adjustments made to fit the theory to the evidence, far exceeding the accommodations (with respect to the imperfection of the fossil record), which Darwin put in place. Nonetheless, the processes advocated by Darwin are independently testable, since he tested his original hypothesis, based on the evidence gathered during the voyage on the Beagle, against evidence gathered after the voyage. Natural selection and common descent prove to be a more parsimonious theory, which has the advantage of explaining bits of evidence problematic to creationists and to allow the same basic principles to be responsible for the diversity of forms across space and time. It is interesting to note how the intertwining and diverse strands of evidence can contribute to the robustness of a theory, although it is also likely that these strands might be in conflict. For instance, paleontological and genetic evidence, might yield contrasting results. In these cases, it is possible that either a revision of the methods employed, of the quality of the record, of the theory under test and of the background assumptions is needed, and the presence of multiple variables at different temporal and spatial scales will make, as argued in § 6, clear cut refutation hard.
- The explanatory and predictive success of Darwin’s theory
In 20th century philosophy of science, the analysis of the relation between hypothesis and evidence generated debate on whether prediction or explanation carried more weight for the successful corroboration of a theory. The predictive successes of the physical sciences had a profound psychological impact on philosophers and scientists. Nonetheless, there are many examples of theories which were successful although involved the explanation of known evidence. Newton’s theory of universal gravitation, for example, provided an explanation of why heavy bodies are attracted to the centre of the Earth, even if this phenomenon was well known to humans in absence of a correct interpretation. Another instance is Harvey’s theory of blood circulation, a process that was accounted for correctly centuries after its discovery. This does not mean that these theories are false and less successful than, say, the prediction of an eclipse. As Peter Achinstein notes, the predictionist and explanationist are committed to understanding the relation of the evidence to the hypothesis in historical terms[27]. Achinstein believes that not always we do need to know what this relation is, and the evaluation of the importance of the evidence depends on other contextual variables. Scriven has noted that in evolutionary biology oftentimes it is easier to explain than to predict certain phenomena, and that in many cases such predictions cannot be easily refuted due to their spatial and temporal inaccuracy[28]. This is due to the presence of factors, such in the case of predicting survival based on fitness, that fall out of the range of accurate predictability: the contingent and historical character of predictions in evolutionary biology highlights the distance from the ‘hard sciences’, although this was not necessarily detrimental to Darwin’s formulation of the theory.
Bromham has drawn on the discussion between the experimental and historical sciences, to argue that it is possible to test predictions at the macro-evolutionary scale[29]. The author takes the term prediction to mean a claim about some unknown fact, whether it happened in the past or in the future. By arguing that laboratory experiments share some of the epistemic issues with ‘natural, real world' experiments, such as the interference of confounding factors and the opacity (i.e. non-linearity) of causal links, Bromham is confident that historical predictions can be tested by building theoretical frameworks that draw on knowledge gained at the micro-evolutionary level, as well as by comparing the outcome of these predictions across multiple lineages. For instance, the hypotheses on mechanisms that brought to development of salt-tolerance in independent plant lineages have been tested and confirmed by cross-comparisons of different taxa; as another instance, the under-determination by different hypotheses of extinction events might be resolved by designing tests which predict an event so singular which corroborates one hypothesis over others. In the case of the extinction of dinosaurs, for example, this kind of evidence might prove useful in determining which explanation is the best. On the hypothesis that a meteorite impact was the main cause of this mass extinctions, there are several factors that make the hypothesis hard to be established. Most fossil discoveries are circumscribed to the North American continent, the margin of error of radiometric dates is broad, and with such restricted evidence it is hard to establish whether this taxon was already in decline world-wide before the impact. In such circumstances, evidential strands, research methods and theories should all cooperate to find the best explanation for this type of events, by broadening the amount and diversity of the evidence gathered and assessing whether the evidence available can be sufficient to confer greater strength to the impact hypothesis over others. The boundary between the explanatory and predictive hypotheses becomes more blurred, although they both turn to be essential in evolutionary biology.
Darwin’s theory employs both explanations and predictions, and I argue that this conjunction allowed the theory to be successful. On the side of explanation, variation in nature and among domestic organisms was well known by breeders, and it was noted how domestic variation was far more limited than that observable in nature. It was commonly held that each species was created independently, and that organisms could not vary indefinitely since they were bound, by God, to stay within the limits of their type. Variation was also contrasted by a tendency to phenotypic characters to blend into each other, and the generation of distinct organism was so hindered.
Darwin had to confront the problem of variation and, although he never understood its causal mechanisms, he thought that the principle of the divergence of characters, contrasting the normalising tendency of natural selection, would have brought to – and hence explained- the origin of new species and of higher taxa[30]. In a similar way, his theory allowed to explain the fossil record – which was already known – but which was accounted for in creationist terms.
At the same time, Darwin made successful and risky predictions. Upon the analysis of the fossil record, he predicted that some ‘proto-bird’ should have marked the transition between reptiles and birds, and this prediction was confirmed with the discovery in 1864 of the Archaeopteryx.
Moreover, he explained and predicted the biogeographical distribution of species living in oceanic islands[31] .
These explanations and predictions show the importance, for Darwin’s theory of a varied methodology. Creationist theories could equally account for the same facts by invoking as many independent acts of creation as needed, and they could only explain the distribution of organisms across space and time, without making any new predictions. There is a sense in which predictions in evolutionary biology might differ from predictions in other sciences. In Darwin’ case, he could make both accurate predictions and more flexible and resilient ones, which did not need extensive detail on the kind and number of organisms at a particular space and time. Nevertheless, even these coarse-grained predictions were relevant to the theory, since they were based on a vast array of evidence, such as migration rates, size and locomotion of the animals and particular features of the plants under consideration, geology and climate of the habitats, and thus constituted an advantage over creationist theories, which were not able to predict anything new. As discussed above, predictions in morphology and embryology were also relevant although perhaps less risky. Furthermore, the success of Darwin’s theoretical insights can be measured by the later applications of the theory. Notwithstanding many predictions turned out to be wrong, variation naturally selected and handed over generations are at the core of contemporary evolutionary biology[32].
Does contemporary evolutionary biology display higher predictive accuracy? It is hard to provide a clear-cut answer. The development of genetics, phylogenetics and paleontological studies seem to have little enhanced the potential of the evolutionary biology in making predictions. At the micro-evolutionary level, it might be easier to predict the outcome of within-population processes in an experimental setting, while in the natural world prediction becomes harder. Recently, predictability has been argued for in the case of micro-organismic evolution[33], and short-term evolution in stick insect populations has been successfully predicted[34]. Although, the contingency of most evolutionary processes makes long-term and fine-grained prediction hard. Nonetheless, there is one field in which some predictions can be attempted: evolutionary ecology[35]. Niles Eldredge has presented the «sloshing-bucket» model accounting for environmental disruptions and subsequent interferences with the extinctions of taxa. Simply put, minor environmental changes would have minor effects on taxa, whilst major changes in the environment could have correspondingly greater effects on the taxa under consideration[36]. It is true that the model was formulated in explaining the paleontological record, although its theoretical structure could be used in predicting the trends of future events. To some extent, then, prediction in evolutionary biology is still possible.
- Does evolutionary biology need any laws to be a science?
The attempt to integrate science with lawfulness has been a major concern of philosophy in the last century. Again, the successes of physics dictated the agenda of those philosophers who were eager to define science in terms of accuracy of predictions and uniformity of nature. This attempt has failed. There is no definition of law of nature agreed upon, universally valid. Humeans, for instance, deny that there is anything describable in terms of nomic necessity, and that physical happenings can be described as accidental regularities, whose inductive recurrence cannot be safely projected into the future[37]. The philosopher Sandra Mitchell has argued for the problematic nature of those approaches that either seek to (a) provide a normative account of laws or to (b) provide paradigmatic laws on which others should be modelled upon. In both cases, the discovery and classification of phenomena as laws is constrained a priori by definitional boundaries, with the risk of excluding important generalisations that do not fit the description[38]. This is more problematic when the source of those normativity and paradigms are the physical sciences. More recently, Tuomas Tahko has defended a hybrid view on the nature of laws, some of which are considered as necessary, while others are contingent. Even this perspective brings to its support case-studies derived from (fundamental) physics and there is a scanty literature considering the presence of laws in evolutionary biology[39]. Elsewhere it has argued for the existence of laws in paleobiology, by formalising them in mathematical language[40]. Turner and Havstad have discussed the merits of Zero-Force Evolutionary Law from a philosophical perspective, arguing that this can allow to cash out trends in macro-evolution in a more regular form: the law states in evolutionary systems, other things being equal, diversity and complexity tend to increase[41]. Nevertheless, Stephen Jay Gould’s studies have had a major influence in shaping the philosophy of biology in emphasising the role of contingence in macro-evolutionary patterns, which might still result from more deterministic and lawlike processes at lower scales[42].
I argue that Darwin’s original theory had succeeded without employing laws or cashing out his theory in a lawful form. Indeed, in The origin ‘law’ is a term that recurs only in relation to variation and ontogenetic development. The Malthusian demographic increase presented in chapter III might be considered as lawful, but it would be probably best described as a trend whose result is contingent on a multiplicity of factors. Furthermore, it is debated whether natural selection is a law. It turns out that the contingence of evolutionary phenomena eludes any attempt to cast evolutionary biology under a lawful form; rather, it is the nature and degree of contingence that set apart biological generalizations from physical laws. The biological world is replete with exceptions to generalisations, and for this reason it is important to emphasise the historical and contextual nature of each explanation. The respectable status of evolutionary biology as a science can be gleaned up from the fact that the core assumptions of Darwin’s theory, descent with modification, are still at the heart of contemporary evolutionary biology and have persisted notwithstanding considerable refinement of the theory.
- The naivety of Popperian crucial experiments
The Popperian emphasis on the possibility of crucial experiments, which would disprove scientific theories notwithstanding its previous support, has been acknowledged to be a simplistic picture of the scientific enterprise[43]. As Imre Lakatos underlined, scientific theories are oftentimes more resilient and scientists do not give up a theory as soon as it is confronted with major issues; rather, they seek to save it[44]. It is indeed reasonable to expect that not all ad hoc accommodations are detrimental to a theory, and Darwin’s case is relevant in this regard. The greatest objection to his theory related to the impossibility of conceiving the usefulness, in an evolutionary perspective, of incipient and not fully formed organs. To this he replied by hypothesising gradual improvements of the organs in question as well as by adopting the idea of functional co-optation, which proposes that the current function of an organ was different from its earlier role. Darwin also had to confront the absence of intermediate stages, which he accounted for in terms of the imperfection of the archaeological record; the loss of certain functions such as sight, to which he provided a Lamarckian explanation, and the ignorance of the causes underlying variation, attempting to explain it through the theory of pangenesis.
All these objections were not enough for Darwin to abandon the theory, and later developments in the 19th and 20th show similar attempts to deal with theoretical issues. This resilience was achieved thanks to the deployment of a plurality of explanatory patterns that stressed the diversity of biological organisms, the need for multiple levels of explanations to account for such differences, which were constituting the protective belt of Darwin’s theory. Rejecting the core of the theory would not have been so straightforward either. Variation was acknowledged to be ubiquitous in nature, and common descent accepted even in a creationist perspective. Many theoreticians were withholding their assent to the principle of natural selection, but it was the methodological and conceptual pluralism of Darwin’s thought as presented in The origin that allowed them to work in a modern, albeit not strictly Darwinian, framework. Moreover, Darwin did not cash out his theory in the formal logic characteristic of 20th century analytic philosophy: identifying a set of assumptions which could be conclusively refuted and bring the theory to the collapse does not do justice to the organic and flexible structure of Darwin’s formulation. Experiments might only be possible at the micro-evolutionary level and no experiment in the narrow sense can be set up at the macro-evolutionary level, where processes occur at large scales in space and time. Even what could be a crucial experiment, or rather observation, for evolutionary theory would not necessarily refute it. If the fossil of a rabbit were to be found in a Precambrian formation, as J.B.S. Haldane stated, that would not be against the whole theory. Evolutionists would be forced to reassess their stratigraphic and phylogenetic theories, but possibly not Darwin’s theory per se. As long as the differential inheritance of some kind of variation occurs, in any form, Darwin’s core theory can be said to remain intact. Moreover, the theory can and does produce statements of a falsifiable character on the logical level, although it is not clear that falsification would result in the outright abandonment of the Darwinian framework; so far the theory has not even been refuted on a metaphysical level. For instance, Lamarckism was at the start of the 20th century a competitor of Darwinism, although it has become clear that it can be integrated – under the form of contemporary epigenetics – rather than opposed to a Darwinian framework.
- Conclusion
In this paper I have sought to defend some theses in the philosophy of science, taking as my starting point the Darwinian theory of evolution as presented in The origin. I have argued that diversity of evidence, the employment of explanations and predictions, and a plurality of patterns at the methodological and conceptual levels are important features of scientific theories – in this case evolutionary biology – and that any monistic interpretation at any of those levels should be resisted. Furthermore, testing those theses against Darwin’s work has allowed to stress the differences between the physical and the biological sciences that, dealing with different entities and processes, require different theoretical frameworks.
Acknowledgments
In structuring this essay and identifying the passages of The origin I have drawn on Telmo Pievani’s work[45].
D. Hume, An enquiry concerning human understanding, Oxford University Press, Oxford 2008.
[2] D. Hull, Darwin’s science and Victorian philosophy of science, in J. Hodge and J. Radick (a cura di), The Cambridge Companion to Darwin, Cambridge University Press, Cambridge 2009, pp. 173-196.
[3] D. Ospovat, The Development of Darwin’s theory, Cambridge University Press, Cambridge 1981.
[4] C.E. Cleland, Historical science, experimental science and the scientific method, in «Geology», XXIX, 11, 2001, pp. 987-990.
[5] A. Currie, Epistemic optimism, speculation and the historical sciences, in «Philosophy, theory and practice in biology», XI, 7, 2019, pp. 1-16.
[6] T. Kuhn, The structure of scientific revolution, Chicago University Press, Chicago 2012.
[7] A. Currie, Marsupial lions and methodological omnivory: function, success and reconstruction in paleobiology, in «Biology and Philosophy», XXX, 2, 2015, pp. 187-209.
[8] S. Toulmin, The uses of argument, Cambridge University Press, Cambridge 2003.
[9]T. Bonnin, Evidential reasoning in the historical sciences: applying Toulmin schemes to the case of Archezoa, in «Biology and Philosophy», XXXIV, 30, 2019, pp. 1-21.
[11] C. Darwin, On the origin of species (1859), Capstone, Padstow 2020, pp. 13-43.
[12] Ibid., pp. XI-XII.
[13] Ibid., p. XII.
[14] Ibid., p. IX.
[15] Ibid., p. X.
[16] C. Hempel, Criteria of confirmation and acceptability, in M. Curd and J.A. Cover (eds.), Philosophy of science: the central issues, Norton, New York 1998, pp. 445-459.
[17] Snell’s law, in optics, exemplifies the relationship between the path taken by a ray of light in crossing the boundary or surface of separation between two contacting substances and the refractive index of each.
[18]J. Kuorikoski, C. Marchionni, Evidential diversity and the triangulation of phenomena, in «Philosophy of Science», LXXXIII, 2, 2016, pp. 227-247.
[19] On empirical adequacy see B. Van Frassen, The scientific image, Clarendon Press, Oxford 1980.
[20] C. Darwin, On the origin of species, cit., pp. 291-292.
[21] Ibid., pp. 292-293.
[22] Ibid., pp. 293-294.
[23] Ibid., pp. 296-320.
[24] This law asserted that «at whatever age any variation first appears in the parent, it tends to reappear at a corresponding age in the offspring» (ibid., p. 372).
[25] Ibid., pp. 368-375.
[26] Ibid., p. XIII.
[27] P. Achinstein, Explanation vs. Prediction: which carries more weight? in M. Curd and J.A. Cover (eds.), Philosophy of science: the central issues, cit., pp. 481-492.
[28] M. Scriven, Explanation and Prediction in evolutionary theory, in «Science», CXXX, 3374, 1959, pp. 477-482.
[29] L. Bromham, Testing hypotheses in macroevolution, in «Studies in the history and philosophy of science», 55, 2016, pp. 47-59.
[30] C. Darwin, On the origin of species, cit., pp. 100-113.
[31] Ibid., pp. 325-331.
[32] T. Pievani, An evolving research programme: The structure of the evolutionary theory in a lakatosian perspective, in A. Fasolo, The theory of evolution and its impact, Springer 2012, pp. 191-212.
[33] A. Mas, Y. Lagadeuc, P. Vandekoornhuyse, Reflections on the predictability of evolution: toward a conceptual framework, in «Science», XXIII, 101736, 2020, pp. 1-11.
[34] P. Nosil, R. Villoutreix, C.F. de Carvalho, T.E. Farkas, V. Soria-Carrasco, J.L. Feder, B.J. Crespi, Z. Gompert, Natural selection and the predictability of evolution in Timema stick insects, in «Science», CCCLIX, 6377, 2018, pp. 765-770.
[35] M. d’Amen, R.G. Mateo, J. Pottier, W. Thuiller, L. Maiorano, L. Pellissier, C. Ndiribe, N. Salamin, A. Guisan, Improving spatial predictions of taxonomic, functional, and phylogenetic diversity, in «Journal of Ecology», CVI, 1, 2018, pp. 76-86.
[36] N. Eldredge, Hierarchies and the sloshing bucket, towards the unification of evolutionary biology, in «Evo. Edu. Outreach», 1, 2008, pp. 10-15.
[37] H. Bhogal, Humeanism about laws of nature, in «Philosophy Compass», XV, 8, 2020, pp. 1-10.
[38] S. Mitchell, Biological complexity and integrative pluralism, Cambridge Studies in the Philosophy of Biology, Cambridge 2003.
[39] T.E. Tahko, The modal status of laws: in defence of a hybrid view, in «The Philosophical Quarterly», LXV, 260, 2015, pp. 509-528.
[40] C.R. Marshall, Five palaeobiological laws needed to understand the evolution of the living biota, in «Nature Ecology and Evolution», I, 6, 2017, pp. 1-6.
[41] D. Turner, C.J. Havstad, Philosophy of macroevolution, Stanford Encyclopedia of Philosophy, 2019, available at: https://plato.stanford.edu/entries/macroevolution/C
[42] S.J. Gould, The structure of evolutionary theory, Belknap University Press, Cambridge-London 2002.
[43] K. Popper, The logic of scientific discovery (1934), Hutchison, London 1959.
[44] I. Lakatos, Falsification and the methodology of scientific research programmes, in J. Worrall, G. Currie (eds.), The Methodology of Scientific Research Programmes, Cambridge University Press, Cambridge 1978, pp. 8-101.
[45] T. Pievani, Anatomia di una rivoluzione, Mimesis, Milano-Udine 2013; Id., Leggere l’origine delle specie di Darwin, Ibis, Como 2015.





